
Abstract
Background&Aims The role of antibody-mediated rejection (ABMR) after liver transplantation (LT) remains controversial. Chronic ABMR (cABMR) is often subclinical and potentially missed without surveillance biopsies (svLbx) which are not established in most LT centers. Transcriptome analysis previously characterized molecular changes in T cell-mediated rejection (TCMR) after solid organ transplantation. We aimed to identify molecular cABMR signatures after LT for a more comprehensive understanding of cABMR.
Methods We retrospectively identified indication and svLbx from our prospective institutional biorepository (n=1207 over 12 years). We performed RNA-sequencing on liver biopsies (discovery cohort: n=71; validation cohort: n=58). Downstream analyses explored the unique and common molecular features of cABMR, clinical (clinTCMR) and subclinical TCMR (subTCMR) compared to no histological rejection (NHR).
Results Nineteen percent of LT recipients with donor-specific antibodies had cABMR. Eighty-one percent of patients with cABMR had ALT and AST ≤2x the upper limit of normal, only being recognized by svLbx. The cABMR group displayed differentially expressed genes (DEGs) uniquely enriched in fibrogenesis-, complement activation- and TNFα-signaling-related pathways. ClinTCMR showed DEGs uniquely enriched in antigen presentation-, interferon-signaling-, and T cell receptor-signaling-related pathways. Common cABMR and clinTCMR DEGs were enriched in chemokine-signaling- and cytokine response-related pathways. Gene set enrichment scores of interferon-signaling and extracellular matrix remodeling pathways discriminated cABMR and clinTCMR. Molecular signatures of clinTCMR correlated with histological TCMR-patterns. Molecular cABMR-signatures correlated with lobular graft injury and liver fibrosis scores. The validation cohort consistently showed patterns of DEGs associating cABMR with fibrogenesis- and NF-κB-signaling-related pathways and clinTCMR with interferon-signaling- and adaptive immunity-related pathways. SubTCMR was molecularly almost indistinguishable from NHR.
Conclusions We report transcriptome-unique features of cABMR that is a unique molecular identity associated with inflammation and fibrogenesis.
Impact and Implications
– Antibody-mediated rejection (ABMR) is a recognized relevant cause of graft injury and unfavorable patient outcomes after kidney, heart and lung transplantation but the role of ABMR after liver transplantation (LT) remains controversial. Hence, we aimed to characterize chronic ABMR after LT on a molecular basis to identify unique features.
– Our findings highlight chronic ABMR to be a distinct clinical phenotype of rejection after LT. Furthermore, we demonstrate chronic ABMR to present mostly with normal liver enzymes, therefore only being detected if protocol biopsies are performed for graft monitoring.
– Therefore, our work emphasizes the usage of protocol graft biopsies for the detection of a fibrosis associated phenotype of cABMR. Pending external validation, the molecular signature may be used in the future to discriminate cABMR from graft injury of other causes.
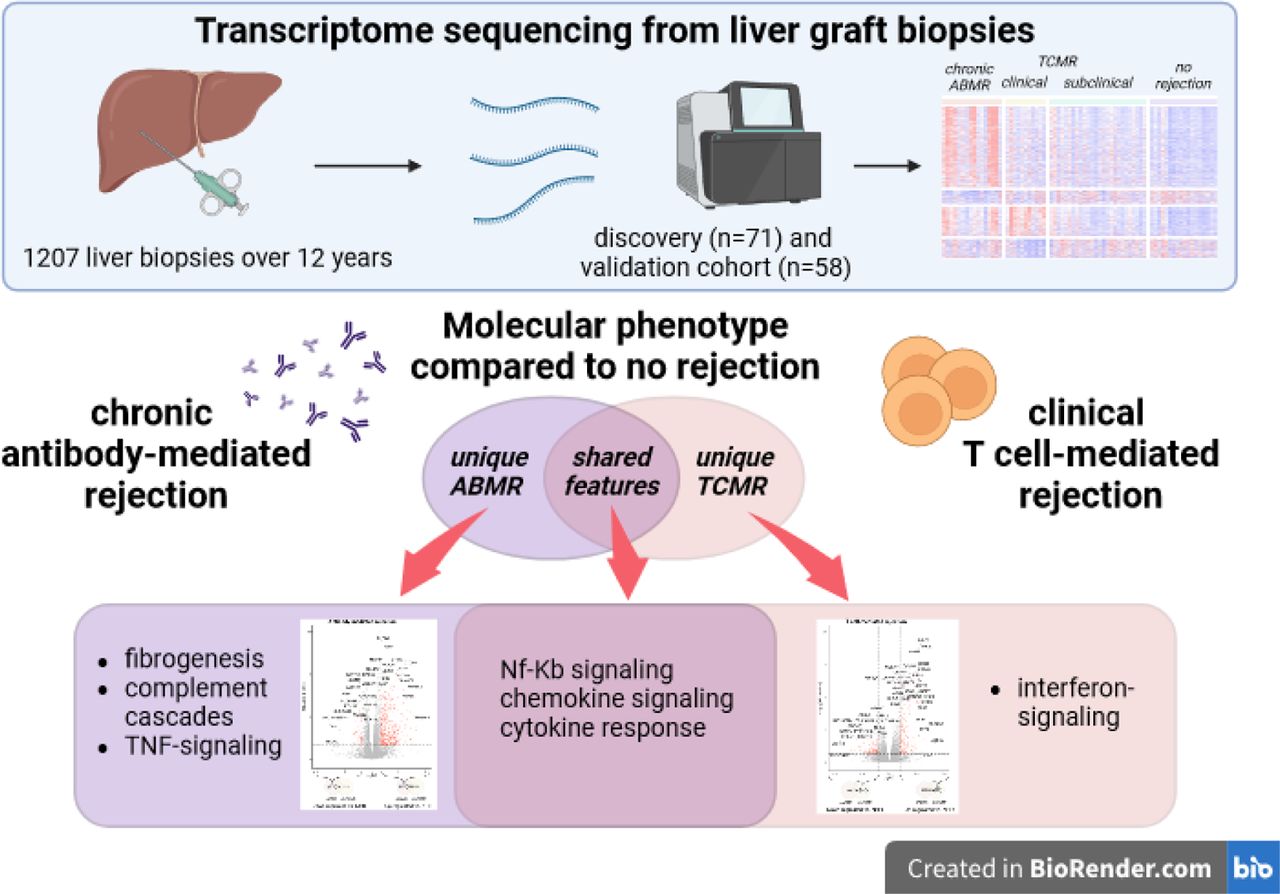
Highlights
– Chronic ABMR was present in up to 1/5 of DSA-positive biopsies and 1/5 of biopsies with advanced fibrosis
– Chronic ABMR and clinically overt TCMR have largely distinct transcriptome patterns with common up-regulated DEGs enriched in pro-inflammatory pathways
– Clinical TCMR is characterized by up-regulated interferon-signaling at the transcriptome level
– Chronic ABMR signatures include up-regulated DEGs enriched in pathways of fibrogenesis, complement cascades, and TNF-signaling
– Common signatures of TCMR and ABMR are enriched in pathways including NF-κB signaling, chemokine signaling and cytokine responses
1. Introduction
Liver transplantation (LT) is a life-saving treatment for patients with end-stage liver disease. Graft survival beyond the first year after LT has not improved in the last decades1. In the long-term, morbidity and mortality are related to both sequelae of immunosuppressive drugs as well as graft dysfunction1. Additionally, surveillance liver biopsy programs reveal advanced fibrosis in 20 - 60% of LT recipients (LTR) after adult and pediatric LT2,3.
The existence and relevance of antibody-mediated rejection (ABMR) in AB0-compatible LT has been a matter of ongoing debate for decades and very few centers are testing for donor-specific anti-HLA antibodies (DSA) regularly as part of their post-transplant follow-up protocols. The presence of DSA is associated with more liver graft injury, more liver graft fibrosis, intrahepatic upregulation of transcripts associated with TCMR, and reduced patient and graft survival2,4–8. The prevalence of DSA increases over time by up to 50% at more than five years after LT2. In 2016, the first consensus document on clinical and histological criteria of acute and chronic ABMR was published by the Banff Foundation, acknowledging a potential role of ABMR after LT but still being controversially discussed9. The low prevalence of acute and chronic ABMR (cABMR) after LT, assumed to be below 10-15%9, prevented faster scientific progress in the understanding of ABMR after LT in comparison to kidney transplantation (KT), where ABMR is one of the major causes of chronic graft dysfunction10. ABMR after LT does not necessarily cause an elevation of liver enzymes above the upper limit of normal (ULN)9. Hence, the prevalence of ABMR is underestimated without quantification of DSA and without surveillance liver graft biopsies (svLBx), both of which are not part of the regular surveillance in the majority of LT centers. Therefore, the true prevalence as well as the implications of cABMR for LTR are unknown.
Gene expression analysis in graft biopsies fostered the characterization of ABMR, especially after KT, and showed comparable molecular signatures of ABMR in kidney, heart and lung transplants11–15, with comparable risk of graft loss in kidney transplant recipients based on their molecular profile irrespective of clinical differences16. Similar gene expression analyses have already identified intrahepatic molecular signatures of spontaneous operational tolerance, of TCMR but, most importantly, not of ABMR17–20. This retrospective single center study aimed to identify molecular signatures of cABMR in distinction to clinically overt TCMR (clinTCMR) and subclinical manifestations of TCMR (subTCMR) in human liver transplant biopsies.
2. Patients and methods
2.1 Patient cohort
Patient samples were retrieved from our ongoing prospective biorepository that was instituted at the end of 2008. The biorepository includes LTR that undergo liver graft biopsy (LBx) either for the work-up of elevated liver enzymes or if signs of liver fibrosis are present, and for surveillance of the liver graft to individualize immunosuppression21. All patients provided written informed consent.
The study was approved by the local Ethics Committee (protocol number 933 for project Z2 of the comprehensive research center 738; MHH Ethikkommission, Hannover, Germany). The study was conducted according to the ethical guidelines of the Declarations of Helsinki and Istanbul.
2.2 Liver biopsies, rejection phenotypes, histological grading and staging
Biopsies of patients with viral or bacterial infection or vascular complications were excluded. Relevant obstructive cholestasis was ruled out by ultrasound examination.
Histological examination and scoring for the rejection activity index (RAI)22, inflammation grade and fibrosis stage (mHAI score)23, central perivenulitis, portal microvasculitis, ductular reaction9, fatty liver disease24 and total liver allograft fibrosis (LAF) score25 was performed by experienced liver pathologists in a blinded fashion. Significant fibrosis was defined as: periportal fibrosis (Ishak F [24]) ≥2 and/or any LAF score component ≥2. SubTCMR and clinTCMR were defined as recently published26.
2.3 RNA isolation and sequencing
For the training cohort, total RNA was extracted with the AllPrep DNA/RNA/Protein Mini kit (ID: 80004, Qiagen N.V., Venlo, The Netherlands) from biopsies freshly preserved in Allprotect Tissue Reagent (ID: 76405, Qiagen N.V., Venlo, The Netherlands) according to the manufacturer’s protocol. For the validation cohort, only FFPE samples were available of which total RNA was extracted with the Qiagen RNeasy FFPE kit (ID: 73504, Qiagen N.V., Venlo, The Netherlands) according to the manufacturer’s protocol. RNA was directly stored at −80 °C until further use. The quality and integrity of the total RNA were assessed using the Agilent Technologies 2100 Bioanalyzer (Agilent Technologies; Waldbronn, Germany) according to the manufacturer’s instructions. Library preparation and sequencing were performed at HZI Braunschweig (Braunschweig, Germany) (training cohort) and BGI Genomics (Hongkong, China) (validation cohort) according to local protocols.
2.4 Gene expression data processing and analysis
Raw sequencing data were firstly processed using nf-core tool (version 3.9) implemented in Nextflow pipelines27,28. Length-scaled gene count tables were imported to Deseq2 (version 1.40.1) using tximport tool (version 1.28.0) for differential expression analysis29,30. Genes were considered to be differentially expressed when the log2 fold change between two conditions exceeded 1, with false-discovery rate (FDR)-adjusted p-value below 0.05. The period in months between the transplantation and the biopsy, recipient age, and sex were included as covariates in the differential gene expression analysis. Functional enrichment of the differentially expressed genes (DEGs) was conducted by the over-representation analysis tool from the ConsensusPathDB-human platform31. For the association between clinical scores and gene expression, the R package limma was used (version 3.56.2)32. The covariates used for the differential expression analysis are also used in the linear regression model to evaluate this association. Gene sets used in gene set enrichment analysis (positive regulation of response to IFNγ (M24135, GO:0060332, version 2023.1.Hs), collagen chain trimeritation (M27812, R-HSA-8948216, version 2023.1.Hs), and laminin interactions (M27216, R-HSA-3000157, version 2023.1.Hs) were downloaded from the GSE broad-institute website33. Methods are outlined further in the supplemental file.
3. Results
3.1 Characterization of the training cohort
The liver biopsies of clinTCMR, subTCMR, cABMR and NHR were selected from a cohort of over 1000 liver biopsies being taken over a period of 12 years. All biopsies with available cryo-conserved tissue were included in the discovery cohort (suppl. Figure 1A). The prevalence of cABMR in DSA positive biopsies was 19% (18/97). Additionally, 19% (18/94) of representative biopsies with advanced fibrosis fulfilled criteria of cABMR.
Patients’ samples were grouped according to their clinical phenotype and the characteristics are outlined in Table 1 and suppl. Figure 2. There were no differences in age between all groups (p=0.30) while sex frequency was different between groups (p=0.04) (Table 1). Time after LT was not different between groups in post-hoc testing (Table 1, suppl. Figure 3A). As per definition, patients with NHR and subTCMR had no elevation of liver enzymes, while these were elevated in patients with clinTCMR. Applying the same threshold of liver enzyme elevation as for clinTCMR and subTCMR, 81% of cABMR had ALT, AST, and AP below two xULN and were therefore categorized as subclinical. This means that they are most likely to be picked up by svLBx.
Histological inflammation and fibrosis were absent in patients with NHR while relevant rejection activity was seen in all other entities being highest in clinTCMR. In contrast, fibrosis according to both Ishak F and LAF scores was highest in cABMR, which also had, by definition, the highest frequency of DSA. Fifty-six percent of biopsies with cABMR also had features of TCMR with an RAI ≥1+1+1, similar as in ABMR after other solid organ transplantation (SOT)34,35. Usage of cyclosporine A (CsA) and tacrolimus (TAC) as primary immunosuppressive drug was different between the groups (p=0.022 and p=0.006 respectively; Table 1). There were no differences regarding the usage of other immunosuppressive agents between the groups.
3.2 Signature genes of cABMR and clinTCMR are associated with distinct biological processes
The study workflow is highlighted in Figure 1A. To investigate the changes in the transcriptome profile in the rejection groups (cABMR, clinTCMR, subTCMR), differential expression analysis of each group compared to NHR was performed in the training cohort (supp. Figure 4; suppl. Table 1). cABMR and clinTCMR shared 369 up-regulated and 54 down-regulated DEGs, while there were 946 up-regulated and 166 down-regulated DEGs unique to cABMR and 354 up-regulated and 223 down-regulated DEGs unique to clinTCMR (Figure 1B; Figure 1C; suppl. Table 3). There were only 35 up-regulated and seven down-regulated DEGs in subTCMR compared to NHR, including two up-regulated and one down-regulated unique DEGs (Figure 1B).

Study design and molecular differences between the rejection groups (A) Study design. (B) Venn diagram showing the numbers of the up-regulated and down-regulated DEGs in each rejection group compared to NHR. (C) Heatmap of scaled gene count. Rows are unique DEGs of cABMR and clinTCMR and columns represent samples. clinTCMR: clinical T cell-mediated rejection; cABMR: chronic antibody-mediated rejection; subTCMR: subclinical T cell-mediated rejection; NHR: no histological rejection, DEGs: differentially expressed genes.
Up-regulated DEGs of cABMR were enriched in processes of fibrogenesis and TNF-signaling (hypergeometric test, FDR-adjusted p <0.05) while up-regulated DEGs of clinTCMR were associated with processes of allograft rejection and cellular responses to cytokines, especially interferon-signaling (hypergeometric test, FDR-adjusted p <0.05) (suppl. Figure 4A, B; suppl. Table 2). Noticeably, the subTCMR transcriptome profile showed minor difference compared to NHR, with up-regulated DEGs involved in antigen presentation, IL-12 production and dendritic cell differentiation (hypergeometric test, FDR-adjusted p <0.05; suppl. Figure 4C; suppl. Table 2).
Next, we explored the transcriptome changes specific to either cABMR or clinTCMR. Unique DEGs in cABMR exhibited enrichment in biological processes of fibrogenesis, cell adhesion and NF-κB-signaling (Figure 2A, B; suppl. Table 4) while unique DEGs in clinTCMR were over-represented in the pathways of allograft rejection, Th1/Th2/Th17 differentiation, antigen processing and presentation, NK cell-mediated cytotoxicity and interferon-signaling among others (Figure 2C, D; suppl. Table 4). To underline the biological relevance of these unique pathways for the pathophysiology of clinTCMR and cABMR, a gene set score enrichment analysis revealed that the positive regulation of response to IFN-gamma was significantly higher in patients with clinTCMR than in patients with cABMR (Student’s t-test, p <0.05; Figure 3A). In contrast, the gene set scores for pathways of restructuration of the basement membrane (“Laminin Interactions”) and the formation of collagen (“Collagen chain trimerization”) were significantly higher in cABMR than in clinTCMR with both processes being relevant to hepatic fibrogenesis (Student’s t-test, p=0.008; Figure 3A).

Molecular signature and functional enrichment of unique DEGs of cABMR and clinTCMR (A) Volcano plot showing the unique cABMR vs NHR DEGs (p-value adjusted by false discovery rate (FDR) <0.05, absolute log2 fold change (LogFc) > 1). (B) Over-represented pathways out of DEGs in panel A. (C) Volcano plot showing the unique clinTCMR vs NHR DEGs (FDR <0.05, LogFc >1) (D) Over-represented pathways out of DEGs in panel C. clinTCMR: clinical T cell-mediated rejection; cABMR: chronic antibody-mediated rejection; subTCMR: subclinical T cell-mediated rejection; NHR: no histological rejection, DEGs: differentially expressed genes.

Gene set enrichment analysis and gene count association with histological scores (A) Gene set scores of three selected pathways in cABMR and clinTCMR. *: p <0.05, **: p <0.01; Student’s t-test. (B) Association between the unique DEGs expression and three different histological scores. Log10(FDR) on the y-axis is signed by the correlation fold change, while DEGs are ranked on the x axis by the association t-stat value. clinTCMR: clinical T cell-mediated rejection; cABMR: chronic antibody-mediated rejection; subTCMR: subclinical T cell-mediated rejection; NHR: no histological rejection, DEGs: differentially expressed genes.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Validation of rejection vs NHR DEGs. DEGs with nominal significance (p <0.05) in the validation cohort are plotted. (B) Over-represented pathways out of DEGs in panel A. clinTCMR: clinical T cell-mediated rejection; cABMR: chronic antibody-mediated rejection; subTCMR: subclinical T cell-mediated rejection; NHR: no histological rejection, DEGs: differentially expressed genes.
The 369 up-regulated DEGs in both clinTCMR and cABMR, highlighted involvement of TNF receptor non-canonical NF-κB-signaling, chemokine-signaling and cytokine response to be essential to both clinTCMR and cABMR (suppl. Figure 5; suppl. Tables 5, 6).
3.3 Association of histopathological scores and differentially expressed genes
Next, we linked unique DEGs to common histopathological scores that assess liver TCMR (RAI), liver allograft fibrosis (LAF) and inflammatory activity (mHAI) (Figure 3B). RAI was most strongly associated with unique DEGs of clinTCMR but only little with unique DEGs of cABMR. DEGs of cABMR correlated most strongly with the LAF score and mHAI, underlining the more lobular graft injury pattern in cABMR. Therefore, histopathological patterns of TCMR (RAI) correlate most strongly with molecular signatures of clinTCMR but less with those of cABMR. In contrast, molecular signatures of cABMR are associated with histopathological patterns of graft fibrosis and lobular graft injury. Thus, clinTCMR and cABMR are characterized by different molecular signatures and different histopathological patterns of injury.
3.4 Validation of possible cABMR and clinTCMR gene signature in an independent validation cohort
The clinical features of the validation cohort were mostly consistent with the training cohort (Table 2). Most notably, the median time after LT at sampling was longer for all phenotypes than in the training cohort, which was accounted for in the biostatistical analysis (see methods section, suppl. Figure 3B). In contrast to the training cohort, primary immunosuppressive therapy was comparable between the groups, because TAC became the leading calcineurin inhibitor over time at our center. Criteria of cABMR were fulfilled in 13% (7/54) of DSA-positive biopsies and 5% (7/135) of biopsies with advanced fibrosis (suppl. Figure 1B). Again, 50% of cABMR had ALT, AST, and AP below two xULN, therefore being likely to be only picked up in svLBx.
Overall consistency of gene expression was high between both cohorts. Eighty-seven percent of cABMR DEGs and 93% of clinTCMR DEGs from the discovery cohort with high-quality RNA from cryoconserved LBx were validated in the validation cohort that used highly degraded RNA being isolated from FFPE tissue (Figure 4A). As in the discovery cohort, up-regulated DEGs in cABMR were enriched in processes of fibrogenesis as well as TNF non-canonical NF-κB-signaling (Figure 4B; suppl. Figure 6A, 7A; suppl. Tables 7, 8). Up-regulated DEGs in clinTCMR were enriched in pathways of Th1/Th2/Th17 cell differentiation, antigen processing and presentation, T cell receptor-signaling, allograft rejection and innate and adaptive immunity (Figure 4B; suppl. Figure 6B; suppl. Tables 7, 8). There were 23 up-regulated and zero down-regulated DEGs in subTCMR compared to NHR, with enrichment in Th1 and Th2 cell differentiation, interferon-signaling, and cellular response to cytokines (suppl. Figure 6C; suppl. Tables 7, 8). DEGs specific to cABMR showed enrichment in the processes of fibrogenesis, cell adhesion, TNFs binding their physiological receptors, TNFR2 non-canonical NF-κB-signaling and complement and coagulation cascades (suppl. Figure 7B; suppl. Tables 9, 10). DEGs specific to clinTCMR were over-represented in pathways of interferon-signaling, antigen processing and presentation, Th1/2/17 cell differentiation, adaptive immunity, T cell receptor-signaling, IL-2-signaling as well as other pathways related to T cells (suppl. Figure 7C; suppl. Tables 9, 10), with findings from the discovery cohort being largely replicated in the validation cohort. Common DEGs of cABMR and clinTCMR were enriched in pathways including cytokine-signaling, especially interferon-gamma, cell adhesion and allograft rejection (suppl. Figure 8A, B; suppl. Tables 11, 12).
4. Discussion
In this study, molecular signatures of three different rejection entities, clinTCMR, cABMR and subTCMR, were characterized using whole-transcriptome sequencing methodologies (mRNA from fresh frozen and FFPE liver biopsies) in two independent cohorts from our center. Hereinafter, we systemically previewed how common and unique features of all three rejection phenotypes highlight potential pathophysiological commonalities and differences. We provide evidence for cABMR to be pathophysiologically distinct from clinTCMR, mainly by fibrogenetic processes, by TNF-signaling via NF-κB as well as complement activation. Lastly, we demonstrated plausibility by linking the molecular phenotype of clinTMCR to hallmark genes of T cell-mediated signaling and to typical histological TCMR patterns by a correlation with the RAI. Likewise, the molecular phenotype of cABMR was associated with genes of fibrogenesis, and histological features of lobular hepatitis and liver fibrosis by a correlation with the HAI and with the LAF score. Of note, in a rat model of ABMR after LT DEGs enriched in pathways of TNF-signaling via NF-κB were more abundant in ABMR36.
Various attempts were made to derive a gene signature of clinTCMR in LTR by using a microarray gene expression approach5,18,19. Most recently, an 11-gene signature showed an excellent diagnostic performance to discriminate clinTCMR from NHR18. Ten out of eleven (all but HLA-DMA with log2 fold change of 0.99 and adjusted p-value of 3.3 x 10-7) of these DEGs were significantly up-regulated (adjusted p-value <0.05 and log2 fold change >1) in clinTCMR compared to NHR in our discovery cohort while 4/11 were up-regulated in the smaller validation cohort (CXCL9, TOP2A, GPNMB, PLA2G7). Of these, only PLA2G7, a gene encoding for a phospholipase that inactivates platelet-activating factor, was uniquely associated with clinTCMR but not with other rejection phenotypes in our study. The low concordance of the 11-gene set with the unique transcriptomic clinTCMR patterns described here underlines the importance of a comprehensive approach including all rejection types for the identification of transcriptomic modules shared by different rejection types and rejection type-specific pathways.
The multicenter INTERLIVER study could not identify ABMR-specific signatures in a cohort of 235 prospectively collected LBx20. This probably relates to the mostly missing DSA testing in the participating centers and the low percentage of svLBx (15%) in this study. The current study selected 22 cABMR cases out of 1207 LBx over a period of 12 years and up to 81% of cABMR had no relevant elevation of liver enzymes and would have been missed without svLBx. So, the missing ABMR signature in the INTERLIVER study potentially relates to a selection bias due to the low frequency of svLBx, again emphasizing the use of svLBx to monitor liver graft function. Of the top ten genes that were most strongly associated with clinTCMR in the INTERLIVER study20, 8/10 (all but PSMB8 and PSMB10) were significantly overexpressed in clinTCMR compared to NHR in our discovery cohort while 6/10 were significantly overexpressed in the validation cohort (GBP1, GBP5, CXCL9, STAT1, PSMB9 and TAP1). Two genes (GBP1 and PSMB9) from the INTERLIVER study were uniquely associated with clinTCMR in our cohort, both of which are interferon-inducible with the former encoding for a guanylate binding protein involved in host defense against pathogens and the latter being related to antigen processing via the immunoproteasome and presentation via MHC-molecules with antigen-recognition by CD8+ T cells37,38. Comparison of published literature with our current study highlights that currently used gene array-based classifiers do not necessarily quantify features unique to clinTCMR but those that may be shared with other rejection phenotypes, e.g. cABMR, in highly-selected, and phenotypically well-characterized cohorts without prevalent ABMR cases. Hence, the final clinical use case of such classifiers will be determined in an unbiased real-world scenario pending further validation. Differences in DEGs between our cohort and previous literature may also be related to different techniques (gene array in the INTERLIVER study and mainly in studies of the Sanchez-Fueyo group18–20,39 with mostly fresh frozen tissue in comparison to whole-transcriptome sequencing in the present study) that were used to quantify gene expression. While it was shown that the number of genes varies between technological platforms, biological processes as well as the performance of diagnostic classifiers were consistent across platforms in patients after KT40. These similar results in terms of biological processes in sequencing techniques from fresh frozen versus FFPE tissue underline that further validation studies can also be performed from stored samples rather than from fresh frozen LBx. Although the BANFF group has recently described the histological changes of cABMR, they are not always clearly applicable in daily clinical practice and are not very specific. Thereby, the diagnosis of cABMR is rarely made based on biopsies. By molecularly identifying patients with cABMR, the diagnostic accuracy of biopsies may be improved.
Concerning cABMR, deposition of complement C4d is regarded as a key feature for the histopathological suspicion of ABMR9. However, it is acknowledged that the performance of various assays to stain C4d is heterogeneous dependent on the technique and the protocol of the local center9. Staining for C4d on FFPE liver tissue exhibits acceptable performance in acute ABMR in comparison to the gold standard of immunofluorescence on frozen tissue, but data on assay performance in cABMR is lacking41. However, it is widely acknowledged that the sensitivity of C4d staining for cABMR is generally low9, 42. Also, C4d deposition may occur in case of biliary obstruction, viral infection of the liver or autoimmune liver injury9 thereby reducing specificity. Given the methodological pitfalls of C4d staining, we included patients with possible cABMR and not only with probable cABMR as defined by the current Banff consensus definition from 20169. Additionally, diagnosing cABMR in the absence of C4d deposition is well established also after KT or lung transplantation with the same prognostic implications as C4d positive ABMR43,44. The consistency of the molecular phenotype found for cABMR in the present study and in published studies from KT supports the validity of our approach45. Hence, our findings fit current pathophysiological concepts of ABMR derived from other SOT, add evidence in favor of the existence of cABMR after LT46.
We report two independent cohorts from our center that are separated by time (discovery cohort: 2008 – 2016, validation cohort 2018 – 2022). In addition, different material with different qualities of RNA was used in these cohorts (discussed above), but we can still demonstrate biological consistency as evidenced by highly overlapping pathways between both cohorts for clinTCMR and cABMR. The molecular profile in terms of biological processes of cABMR was largely maintained in the validation cohort, despite lower sample numbers for cABMR. Nonetheless, these findings have to be validated externally by future multicenter efforts. However, a reasonable amount of cABMR may be missed in such studies if svLBx and DSA testing were not performed routinely as up to 81% of cABMR in our cohort had liver enzymes below two xULN and would not be eligible for liver biopsy in studies that do not incorporate svLBx47,48.
We highlight biological processes of cABMR taking place in LTR that may provide druggable targets in the future to prevent disease progression. Concerning the latter, implementation of svLBx programs3,21 will probably be a relevant prerequisite to determine the real prevalence of cABMR across countries, centers, and IS regimens. SvLBx programs reported superior survival of participants3. These programs select long-term survivors, without contraindications for a LBx and with a high adherence and motivation to perform an invasive measurement to potentially decrease their long-term maintenance immunosuppression. The broader application of DSA measurement and elastography for the assessment of graft fibrosis can help to enrich for LTR with relevant graft injury 2,49,50. In case of relevant fibrosis or positivity for DSA, a biopsy is often warranted.
This study indicates cABMR after LT as a unique entity with a similar gene signature as described in other SOT. The distinct phenotype of cABMR after LT is characterized by fibrogenesis, and TNF-mediated signaling via NF-κB and involves endothelial cell-signaling as well as complement cascades. The cABMR gene signature is closely associated with histological fibrosis (LAF score) and is frequently detected in DSA-positive LTR. Pending external validation, we highlight a possible application of RNA-sequencing on liver biopsies to facilitate the detection of cABMR in LTR.
Data Availability
The data that support the plots within this paper and other findings of this study are available from the corresponding author upon reasonable request. The raw gene expression data, including a sample-data relationship file, as well as the count tables are stored conformant with the MINSEQE reporting standard (Version 1.0, June 2012, accessible at https://zenodo.org/doi/10.5281/zenodo.5706411) at MHH RepoMed repository https://doi.org/10.26068/mhhrpm/20240207-000.
Acknowledgements
We thank Szilvia Ziegert and Konstantinos Iordanidis from the “immune tolerance working group” at Hannover Medical School for technical assistance. We thank Stefan Huebscher, already retired from his work in the pathology department of the University of Birmingham/UK, for his advice in initiating this project and for his training of the pathologists at Hannover Medical School during a fruitful workshop in September 2016.
Footnotes
↵* authors share first authorship
↵+ authors share last authorship
↵11 Biomedical Research in End-stage and Obstructive Lung Disease Hannover (BREATH) of the German Center for Lung Research (DZL), Hannover, Germany
Financial support: The work was supported by grants from the German Research Foundation (SFB738 project Z2; EJ)), the Transplantation Center Project 19_02 and HiLF II from Hannover Medical School (RT) and the Transplantation Center Project ZN3369 from Hannover Medical School/The Ministry of Science and Culture of the State of Lower Saxony (BE). RT was supported by the Core 100 advanced clinician scientist program from Hannover Medical School. BE was supported by the PRACTIS – Clinician Scientist program of Hannover Medical School, funded by the German Research Foundation (DFG, ME 3696/3). ACM was supported by the DAAD Research Grants – One-year grants for doctoral Candidates, 2023/2024 (57645447). KC was supported by the Else Kröner-Fresenius Stiftung (KlinStrucMed programme). This work was supported by the Radboud University Medical Centre Hypatia Grant, an ERC Starting Grant, grant agreement 948207 (ModVaccine), the Deutsche Forschungsgemeinschaft (DFG, German Research Foundation) under Germany’s Excellence Strategy - EXC 2155 - project number 390874280, and the Lower Saxony Center for AI and Causal Methods in Medicine (CAIMed) grant funded by the Ministry of Science and Culture of Lower Saxony with funds from the program zukunft.niedersachsen of the VolkswagenStiftung (all to YL).
Conflict of interest: Nothing to declare with regard to this manuscript.
Data availability statement: The data that support the plots within this paper and other findings of this study are available from the corresponding author upon reasonable request. The raw gene expression data, including a sample-data relationship file, as well as the count tables are stored conformant with the MINSEQE reporting standard (Version 1.0, June 2012, accessible at https://zenodo.org/doi/10.5281/zenodo.5706411) at MHH RepoMed repository, https://doi.org/10.26068/mhhrpm/20240207-000.
Abbreviations
- ABMR
- antibody-mediated rejection
- cABMR
- chronic antibody-mediated rejection
- CsA
- cyclosporine A
- clinTCMR
- clinically overt T cell-mediated rejection
- DEGs
- differentially expressed genes
- DSA
- donor-specific anti-HLA antibodies
- FDR
- false-discovery rate
- FFPE
- formalin-fixed paraffin-embedded
- KT
- kidney transplantation
- LAF
- liver allograft fibrosis
- LBx
- liver graft biopsy
- LT
- liver transplantation
- LTR
- liver transplant recipient
- NHR
- no histological rejection
- mHAI
- modified histological activity index
- RAI
- rejection activity index
- RIN
- RNA integrity number
- SOT
- solid organ transplantation
- subTCMR
- subclinical T cell-mediated rejection
- svLBx
- surveillance liver graft biopsies
- TAC
- tacrolimus
- TCMR
- T cell-mediated rejection
- ULN
- upper limit of normal
- x ULN
- times the upper limit of normal.
References




